Splicing des exitrons : Le mécanisme négligé révolutionnant notre compréhension de l’expression des gènes. Découvrez comment cet événement de splicing alternatif façonne la complexité du protéome et la maladie.
- Introduction au Splicing des Exitron
- Découverte Historique et Nomenclature
- Mécanismes Moléculaires Sous-jacents au Splicing des Exitron
- Approches Bioinformatiques pour la Détection des Exitron
- Conséquences Fonctionnelles sur la Structure Protéique
- Splicing des Exitron dans la Santé et la Maladie
- Analyse Comparative à Travers les Espèces
- Méthodes Expérimentales pour la Validation
- Implications Thérapeutiques et Biotechnologiques
- Orientations Futures et Questions Ouvertes
- Sources & Références
Introduction au Splicing des Exitron
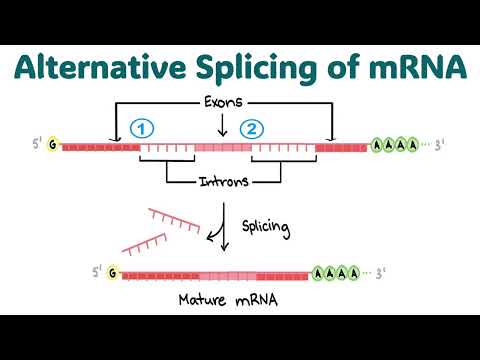
Le splicing des exitrons est un phénomène de splicing alternatif dans lequel les régions internes des exons codants pour des protéines, appelées « exitrons », sont supprimées de manière sélective des transcrits de pré-ARNm. Contrairement aux introns canoniques, les exitrons sont intégrés dans des exons annotés et leur excision peut conduire à la production de divers isoformes de protéines ayant une structure et des fonctions modifiées. Ce processus élargit la complexité protéomique des organismes eucaryotes et a des implications significatives pour la régulation des gènes, l’adaptation cellulaire et la pathogénie des maladies.
Le terme « exitron » a été introduit pour la première fois pour décrire les séquences exoniques qui se comportent de manière similaire aux introns, étant épissées dans certaines conditions. Le splicing des exitrons est distinct du saut d’exon traditionnel ou de la rétention d’intron, car il implique l’élimination de séquences qui sont généralement considérées comme faisant partie de la région codante. L’ARNm résultant peut coder des protéines avec des délétions internes, affectant potentiellement des domaines critiques pour l’activité, la localisation ou les interactions des protéines.
Des avancées récentes dans le séquençage d’ARN à haut débit et l’analyse computationnelle ont permis l’identification systématique des événements de splicing des exitrons à travers diverses espèces, y compris les humains, les plantes et les organismes modèles. Ces études ont révélé que le splicing des exitrons est un mécanisme répandu et conservé, contribuant à la diversité transcriptomique et protéomique. Notamment, le splicing des exitrons a été impliqué dans la régulation de processus biologiques clés tels que la différenciation cellulaire, les réponses au stress et la fonction immunitaire.
Les conséquences fonctionnelles du splicing des exitrons dépendent du contexte. Dans certains cas, l’élimination d’un exitron peut générer des isoformes protéiques avec des fonctions nouvelles ou dominantes-négatives, tandis que dans d’autres, cela peut mener à la production de protéines tronquées ou non fonctionnelles. La dérégulation du splicing des exitrons a été associée à diverses maladies, y compris le cancer, où des modèles de splicing aberrants peuvent conduire à la tumorigenèse ou influencer la résistance thérapeutique. Comprendre les mécanismes régissant la reconnaissance et l’excision des exitrons est donc d’un intérêt considérable tant pour la biologie fondamentale que pour la recherche clinique.
La recherche sur le splicing des exitrons est soutenue par des organisations scientifiques importantes et des instituts de recherche dans le monde entier, y compris les National Institutes of Health et l’European Bioinformatics Institute, qui fournissent des ressources et des bases de données pour l’étude des événements de splicing alternatif. À mesure que le domaine progresse, élucider les réseaux régulateurs et les résultats fonctionnels du splicing des exitrons sera crucial pour tirer parti de son potentiel dans le diagnostic et les thérapeutiques.
Découverte Historique et Nomenclature
Le splicing des exitrons représente une addition relativement récente au paysage en expansion des événements de splicing alternatif dans les transcriptomes eucaryotes. Le terme « exitron » est un mot-valise de « intron exoniques », reflétant la nature unique de ces séquences : ce sont des régions de type intron intégrées dans des exons codants pour des protéines annotés. La découverte historique du splicing des exitrons peut être retracée aux avancées des technologies de séquençage RNA à haut débit (RNA-seq) au début des années 2010, qui ont permis aux chercheurs de détecter des événements de splicing auparavant non reconnus à une résolution de nucléotides uniques.
La première identification systématique et caractérisation du splicing des exitrons a été rapportée en 2014 par Marquez et al., qui ont analysé les transcriptomes d’Arabidopsis thaliana et de cellules humaines. Leur travail a révélé que certaines régions exoniques pouvaient être alternativement épissées, se comportant de manière similaire aux introns conventionnels mais résidant dans des exons annotés. Cette découverte a remis en question la classification binaire traditionnelle des exons et des introns, suggérant une vision plus nuancée de l’architecture des gènes et de la diversité des transcrits. Les auteurs ont inventé le terme « exitron » pour décrire ces régions, en mettant l’accent sur leurs caractéristiques exoniques et introniques duales.
La nomenclature « exitron » a depuis été largement adoptée dans la littérature scientifique pour distinguer ces éléments des introns et des exons canoniques. Le splicing des exitrons est maintenant reconnu comme un mécanisme conservé à travers diverses lignées eucaryotes, y compris les plantes, les animaux et les champignons. La découverte des exitrons a incité à une réévaluation des pratiques d’annotation des gènes et a mis en lumière la complexité de la régulation post-transcriptionnelle. Notamment, le splicing des exitrons peut générer des isoformes protéiques avec des structures de domaines modifiées, affectant potentiellement la fonction protéique et les phénotypes cellulaires.
L’intérêt croissant pour le splicing des exitrons a conduit au développement d’outils computationnels spécialisés et de bases de données pour leur identification et annotation. Les principales organisations de recherche et les consortiums génomiques, tels que l’European Bioinformatics Institute et le National Center for Biotechnology Information, ont intégré des données liées aux exitrons dans leurs ressources, facilitant l’exploration de ce phénomène. À mesure que le domaine continue d’évoluer, la découverte historique et la nomenclature du splicing des exitrons soulignent la nature dynamique de la biologie du génome et le raffinement continu de notre compréhension de la régulation de l’expression des gènes.
Mécanismes Moléculaires Sous-jacents au Splicing des Exitron
Le splicing des exitrons est une forme récemment caractérisée de splicing alternatif qui implique l’excision de séquences codantes internes, appelées « exitrons », à partir des transcrits d’ARNm matures. Contrairement aux introns canoniques, les exitrons sont situés dans des exons codants pour des protéines annotés, et leur élimination peut modifier de manière significative le potentiel codant de l’ARNm résultant. Les mécanismes moléculaires sous-jacents au splicing des exitrons sont complexes et impliquent à la fois des éléments cis-régulateurs et des facteurs de splicing trans-agissants.
Au cœur du splicing des exitrons se trouve la reconnaissance de sites de splice non canoniques au sein des régions exoniques. Les exitrons possèdent généralement des séquences consensus de site de splice plus faibles par rapport aux introns conventionnels, ce qui rend leur reconnaissance par le spliceosome moins efficace et plus dépendante du contexte. Le spliceosome, un complexe ribonucléoprotéique dynamique responsable du splicing de l’ARNm précurseur, doit distinguer les limites des exitrons des séquences exoniques environnantes. Ce processus est influencé par la présence d’activateurs de splicing exoniques (ESE) et de silences (ESS), qui recrutent ou repoussent des protéines riches en sérine/arginine (SR) et des ribonucléoprotéines nucléaires hétérogènes (hnRNP) qui modulent la sélection des sites de splice.
Les facteurs trans-agissants jouent un rôle essentiel dans la régulation du splicing des exitrons. Les protéines SR favorisent généralement la reconnaissance des sites de splice et l’inclusion des séquences exoniques, tandis que les hnRNP agissent souvent comme récepteurs, privilégiant l’excision des exitrons. L’équilibre entre ces facteurs, ainsi que leurs niveaux d’expression et leurs modifications post-traductionnelles, peut faire basculer le résultat du splicing. De plus, l’environnement chromatinien local et les taux d’élongation de l’ARN polymérase II ont été montrés pour influencer les décisions de splicing alternatif, y compris l’utilisation des exitrons, en modulant l’accessibilité des machines de splicing aux transcrits naissants.
Des études récentes ont souligné la conservation évolutive du splicing des exitrons à travers les eucaryotes, suggérant un rôle biologique fondamental. Dans les plantes, par exemple, le splicing des exitrons a été impliqué dans la diversification du protéome et les réponses au stress, tandis que chez les humains, il est de plus en plus reconnu comme une source de diversité transcriptomique et protéomique, avec des implications potentielles dans le cancer et d’autres maladies. Les conséquences fonctionnelles du splicing des exitrons sont diverses, allant de la génération de nouvelles isoformes protéiques à l’introduction de codons d’arrêt prématurés, ce qui peut déclencher la dégradation médiée par des erreurs.
La recherche en cours, soutenue par des organisations telles que les National Institutes of Health et l’European Bioinformatics Institute, continue d’élucider les déterminants moléculaires précis et les réseaux régulateurs gouvernant le splicing des exitrons. Les avancées dans le séquençage à haut débit et l’analyse computationnelle devraient encore défaire la complexité de ce mécanisme de splicing alternatif et son impact sur la régulation de l’expression des gènes.
Approches Bioinformatiques pour la Détection des Exitron
Le splicing des exitrons représente une forme non canonique de splicing alternatif, où des régions internes des exons codants annotés – appelées « exitrons » – sont excisées de l’ARNm mature. Ce processus peut générer des isoformes protéiques avec des fonctions modifiées et a été impliqué à la fois dans la physiologie normale et dans les maladies, y compris le cancer. La détection des événements de splicing des exitrons pose des défis bioinformatiques uniques, car les exitrons ne sont pas annotés comme des introns conventionnels et leur splicing peut dépendre du contexte. Par conséquent, des approches computationnelles spécialisées ont été développées pour identifier et caractériser avec précision le splicing des exitrons à partir de données de séquençage RNA à haut débit (RNA-seq).
L’étape principale dans la détection des exitrons implique l’alignement des lectures de RNA-seq à un génome ou transcriptome de référence. Des outils d’alignement standard tels que STAR et HISAT2, développés par le National Center for Biotechnology Information et d’autres consortiums de recherche, sont couramment utilisés à cet effet. Cependant, parce que les exitrons sont intégrés dans des exons, les aligners conscients du splicing traditionnels peuvent ne pas toujours faire la distinction entre le splicing des exitrons et les jonctions exon-exon canoniques. Pour y remédier, des outils dédiés tels que « ScanExitron » et « Exitron-Seq » ont été développés. Ces outils tirent parti des signatures de séquence uniques du splicing des exitrons – spécifiquement, la présence de jonctions de splice non canoniques au sein des exons annotés – pour identifier les événements de splicing des exitrons candidats.
Les pipelines bioinformatiques pour la détection des exitrons incluent généralement plusieurs étapes clés :
- Alignement des Lectures : Cartographie de haute qualité des lectures de RNA-seq au génome de référence, en prêtant attention aux lectures éclatées qui peuvent indiquer de nouvelles jonctions de splice au sein des exons.
- Identification des Jonctions : Extraction des jonctions de splice à partir des fichiers d’alignement, en se concentrant sur celles qui ne correspondent pas aux frontières intron-exon annotées.
- Filtrage des Candidats Exitron : Application de filtres pour distinguer les véritables événements de splicing des exitrons des artefacts de séquençage ou des mauvais alignements, utilisant souvent des critères tels que le soutien minimal en lectures, les motifs de sites de splice canoniques, et la conservation à travers des échantillons.
- Annotation et Quantification : Intégration avec des bases de données d’annotation de gènes, telles que celles maintenues par Ensembl ou GENCODE, pour mapper les événements de splicing des exitrons à des gènes spécifiques et quantifier leur utilisation à travers des conditions.
Des avancées récentes dans les technologies de séquençage à longues lectures, soutenues par des organisations comme Pacific Biosciences et Oxford Nanopore Technologies, ont encore amélioré la détection des exitrons en permettant l’observation directe de transcrits complets et de modèles de splicing complexes. Ces technologies réduisent l’ambiguïté dans l’attribution des jonctions de splice et facilitent la découverte d’événements de splicing des exitrons nouveaux qui peuvent être manqués par des approches à courtes lectures.
En résumé, la détection bioinformatiques du splicing des exitrons repose sur une combinaison d’algorithmes d’alignement avancés, d’outils de détection spécialisés et d’une intégration avec des ressources complètes d’annotation des gènes. À mesure que les technologies de séquençage et les méthodes computationnelles continuent d’évoluer, la sensibilité et la spécificité de la détection des exitrons devraient s’améliorer, approfondissant notre compréhension de ce phénomène de splicing intrigant.
Conséquences Fonctionnelles sur la Structure Protéique
Le splicing des exitrons est une forme de splicing alternatif dans laquelle des régions internes des exons codants pour des protéines, appelées « exitrons », sont excisées des transcrits de pré-ARNm. Ce processus peut avoir des conséquences fonctionnelles profondes sur la structure de la protéine résultante, car il altère directement la séquence d’acides aminés codée par les exons affectés. Contrairement aux introns canoniques, les exitrons sont intégrés dans des séquences exoniques et leur élimination ne perturbe pas le cadre de lecture dans la plupart des cas, mais peut conduire à la production d’isoformes protéiques avec des domaines, des motifs ou des sites fonctionnels modifiés.
L’excision des exitrons peut entraîner la suppression de segments protéiques spécifiques, supprimant ou modifiant potentiellement des domaines fonctionnels tels que des sites catalytiques actifs, des motifs de liaison ou des régions régulatrices. Cela peut influencer la stabilité, la localisation de la protéine, son interaction avec d’autres molécules et son activité biologique globale. Par exemple, si un exitron code une partie d’un domaine catalytique, son élimination peut rendre la protéine enzymatiquement inactive ou altérer sa spécificité de substrat. À l’inverse, la rétention d’un exitron peut préserver ces domaines, conduisant à l’expression de l’isoforme protéique canonique.
Des études structurelles ont montré que le splicing des exitrons peut engendrer des variantes protéiques avec des conformations tridimensionnelles distinctes. Ces changements structurels peuvent influencer le repliement des protéines, l’oligomérisation, ou la capacité de former des complexes avec d’autres protéines ou acides nucléiques. Dans certains cas, le splicing des exitrons introduit de nouvelles jonctions de séquence qui peuvent créer de nouveaux épitopes ou sites de modification post-traductionnelle, diversifiant encore le protéome. L’impact fonctionnel de ces changements dépend du contexte et peut varier d’une modulation subtile de l’activité à une perte ou un gain complet de fonction.
Il est important de noter que le splicing des exitrons n’est pas aléatoire mais est régulé de manière spécifique aux tissus et au développement, suggérant qu’il joue un rôle dans l’affinement de la fonction des protéines en réponse aux besoins physiologiques. La dérégulation du splicing des exitrons a été impliquée dans diverses maladies, y compris le cancer, où le splicing aberrant peut conduire à la production de variantes protéiques oncogènes ou à la perte des fonctions des suppresseurs de tumeur. L’étude du splicing des exitrons et de ses effets sur la structure protéique est un domaine de recherche actif, avec des implications pour comprendre la complexité du protéome et développer des stratégies thérapeutiques ciblées.
La recherche sur les mécanismes et les conséquences du splicing des exitrons est soutenue par des organisations scientifiques de pointe telles que les National Institutes of Health et l’European Bioinformatics Institute, qui fournissent des ressources et des bases de données pour l’analyse des événements de splicing alternatif et leur impact sur la structure et la fonction des protéines.
Splicing des Exitron dans la Santé et la Maladie
Le splicing des exitrons est une forme récemment caractérisée de splicing alternatif qui implique l’excision de séquences codantes internes, appelées « exitrons », à partir d’exons annotés codants pour des protéines. Contrairement aux introns canoniques, les exitrons sont intégrés dans des exons et leur élimination ou rétention peut considérablement modifier le produit protéique résultant. Ce processus augmente la diversité protéomique et la complexité fonctionnelle des cellules eucaryotes, avec des implications significatives pour la physiologie normale et les états pathologiques.
Dans les tissus sains, le splicing des exitrons contribue à l’ajustement de l’expression des gènes et de la fonction des protéines. En générant plusieurs isoformes de protéines à partir d’un seul gène, le splicing des exitrons permet aux cellules de s’adapter aux signaux de développement et aux changements environnementaux. Par exemple, chez les plantes, le splicing des exitrons a été montré comme jouant un rôle dans les réponses au stress et la régulation du développement, comme documenté par des recherches menées par l’European Bioinformatics Institute (EMBL-EBI). Chez les humains, le splicing des exitrons est de plus en plus reconnu comme un mécanisme destiné à élargir le répertoire fonctionnel des protéines, en particulier dans des tissus avec une grande diversité cellulaire tels que le cerveau et le système immunitaire.
Cependant, la dérégulation du splicing des exitrons a été impliquée dans diverses maladies, notamment le cancer. Le splicing aberrant des exitrons peut conduire à la production de protéines tronquées ou modifiées qui peuvent favoriser l’oncogenèse ou conférer une résistance à la thérapie. Par exemple, des études ont identifié des événements de splicing des exitrons récurrents dans des gènes associés à la suppression des tumeurs et à la régulation du cycle cellulaire, suggérant un rôle dans la progression tumorale. Le National Cancer Institute souligne l’importance du splicing alternatif, y compris les événements d’exitrons, dans la génération de néo-antigènes susceptibles d’être reconnus par le système immunitaire, offrant ainsi des cibles potentielles pour l’immunothérapie.
Au-delà du cancer, le splicing des exitrons a été lié à des troubles neurodégénératifs et à des maladies génétiques. La mauvaise épissure des exitrons dans des gènes neuronaux peut perturber la fonction synaptique et contribuer à des conditions telles que la sclérose latérale amyotrophique (SLA) et certaines formes d’épilepsie. Les National Institutes of Health soutiennent les recherches en cours sur les mécanismes moléculaires sous-jacents au splicing des exitrons et leur impact sur la santé humaine.
À mesure que les technologies de séquençage à haut débit et les outils computationnels avancent, le paysage du splicing des exitrons dans la santé et la maladie devient plus clair. Comprendre les réseaux régulateurs et les conséquences fonctionnelles du splicing des exitrons ouvre la voie au développement de nouveaux marqueurs diagnostiques et de stratégies thérapeutiques, soulignant son importance dans la médecine moléculaire.
Analyse Comparative à Travers les Espèces
Le splicing des exitrons, une forme de splicing alternatif où des régions exoniques internes (exitrons) sont excisées de l’ARNm mature, a émergé comme un mécanisme significatif pour élargir la diversité transcriptomique et protéomique chez les eucaryotes. Les analyses comparatives à travers les espèces révèlent à la fois des caractéristiques conservées et divergentes du splicing des exitrons, mettant en évidence son importance évolutive et fonctionnelle.
Chez les plantes, le splicing des exitrons a d’abord été systématiquement caractérisé dans Arabidopsis thaliana, où il a été montré qu’il contribue à la complexité du protéome en générant des isoformes protéiques avec des domaines ou des motifs régulateurs modifiés. Des études menées par l’Arabidopsis Information Resource ont catalogué de nombreux événements de splicing des exitrons, démontrant leur prévalence et leurs rôles potentiels dans les réponses au stress et le développement. Notamment, les exitrons des plantes conservent souvent un potentiel de codage, et leur splicing est étroitement régulé en réponse à des signaux environnementaux.
Chez les animaux, le splicing des exitrons a été observé dans divers taxa, y compris les mammifères, les insectes et les nématodes. Chez les humains, des recherches soutenues par le National Center for Biotechnology Information (NCBI) et les National Institutes of Health (NIH) ont identifié des événements d’exitrons dans des tissus normaux et cancéreux. Les exitrons humains se chevauchent souvent avec des exons codants pour des protéines, et leur excision peut entraîner des décalages de cadre, des codons d’arrêt prématurés, ou la suppression de domaines protéiques fonctionnels. Cela a des implications pour la maladie, en particulier dans l’oncogenèse, où un splicing aberrant des exitrons peut générer des néo-antigènes ou perturber des gènes suppresseurs de tumeurs.
Les analyses de génomique comparative indiquent que bien que le mécanisme de base du splicing des exitrons soit conservé, la fréquence, les éléments régulateurs et les conséquences fonctionnelles varient entre les espèces. Par exemple, le splicing des exitrons semble plus répandu chez les plantes que chez les animaux, ce qui pourrait refléter des différences dans l’organisation du génome et les machines de splicing. La base de données génomique Ensembl, maintenue par l’European Bioinformatics Institute, fournit des annotations inter-espèces qui facilitent de telles études comparatives, révélant des modèles spécifiques aux lignées et la conservation évolutive des gènes contenant des exitrons.
De plus, les facteurs régulateurs gouvernant le splicing des exitrons, tels que la force des sites de splice et la présence de protéines liantes de l’ARN spécifiques, présentent des caractéristiques à la fois conservées et spécifiques aux espèces. La recherche en cours, soutenue par des organisations telles que l’European Molecular Biology Laboratory (EMBL), continue d’élucider les déterminants moléculaires et les conséquences biologiques du splicing des exitrons à travers l’arbre de la vie.
Méthodes Expérimentales pour la Validation
La validation expérimentale du splicing des exitrons est essentielle pour confirmer les prédictions computationnelles et élucider la signification biologique de ces événements de splicing non canonique. Le splicing des exitrons, qui implique l’excision de séquences codantes internes (régions exitroniques) des ARN matures, peut être validé en utilisant une combinaison de techniques de biologie moléculaire, de séquençage à haut débit et d’essais fonctionnels.
Une approche fondamentale pour valider le splicing des exitrons est la réaction en chaîne par polymérase de transcription inverse (RT-PCR). Les chercheurs conçoivent des amorces encadrant la région d’exon prédite pour amplifier à la fois les isoformes épissées et non épissées à partir de l’ADN complémentaire (ADNc) dérivé d’échantillons d’ARN. La présence de produits PCR distincts correspondant à l’inclusion ou à l’exclusion de l’exitron peut être visualisée par électrophorèse sur gel. Le séquençage Sanger de ces produits confirme en outre les jonctions de splice précises, fournissant des preuves directes du splicing des exitrons au niveau du transcript.
La PCR quantitative en temps réel (qRT-PCR) est souvent utilisée pour mesurer l’abondance relative des transcrits spliced par rapport aux transcrits canoniques. Cette méthode permet d’évaluer la fréquence du splicing des exitrons à travers différents tissus, stades de développement ou conditions expérimentales. Pour une résolution et un débit plus élevés, le séquençage RNA (RNA-seq) est largement utilisé. En cartographiant les lectures de séquençage au génome et au transcriptome de référence, les chercheurs peuvent identifier des lectures qui s’étendent sur de nouvelles jonctions de splice indiquant l’excision des exitrons. Des outils computationnels spécifiquement conçus pour la détection des exitrons, tels que ceux tirant parti des alignements de lectures éclatées, améliorent la sensibilité et la spécificité de la validation basée sur le RNA-seq.
Pour confirmer que le splicing des exitrons conduit à la production d’isoformes protéiques modifiées, la protéomique basée sur la spectrométrie de masse peut être utilisée. Cette approche détecte des peptides uniques aux isoformes splicing des exitrons, fournissant des preuves directes au niveau protéique. De plus, le western blotting avec des anticorps spécifiques aux isoformes peut valider l’expression des protéines résultant du splicing des exitrons.
La validation fonctionnelle implique souvent l’utilisation d’essais de rapporteurs de minigène. Dans cette méthode, des fragments génomiques contenant l’exitron et ses exons flanquants sont clonés dans des vecteurs d’expression et transfectés dans des cellules cultivées. Le modèle d’épissage du transcript du minigène est ensuite analysé par RT-PCR ou séquençage, permettant aux chercheurs de décomposer les éléments cis-régulateurs et les facteurs trans-agissants influençant le splicing des exitrons.
Dans l’ensemble, ces méthodes expérimentales – allant de la RT-PCR et du RNA-seq à la protéomique et aux essais de minigène – fournissent une boîte à outils complète pour valider les événements de splicing des exitrons et étudier leurs conséquences fonctionnelles. Ces approches sont largement adoptées et recommandées par les principales organisations de recherche telles que les National Institutes of Health et l’European Bioinformatics Institute, qui soutiennent le développement et la diffusion de bonnes pratiques dans la recherche en biologie de l’ARN.
Implications Thérapeutiques et Biotechnologiques
Le splicing des exitrons, une forme de splicing alternatif où des régions exoniques internes (exitrons) sont supprimées de manière sélective de l’ARNm mature, a émergé comme un mécanisme significatif influençant la diversité protéomique et la régulation des gènes. La découverte du splicing des exitrons a de profondes implications thérapeutiques et biotechnologiques, en particulier dans le contexte des maladies humaines et de la biologie synthétique.
En oncologie, le splicing des exitrons a montré qu’il génère des isoformes protéiques nouvelles qui peuvent contribuer à la tumorigenèse, à l’évasion immunitaire et à la résistance aux médicaments. Par exemple, les événements de splicing aberrants d’exitrons peuvent produire des protéines tronquées ou altérées qui favorisent la progression du cancer ou créent des néo-antigènes reconnaissables par le système immunitaire. Cela ouvre des voies pour le développement d’immunothérapies anticancéreuses ciblant des néoépitopes dérivés des exitrons, ainsi que de petites molécules ou d’oligonucléotides antisens conçus pour moduler les modèles de splicing des exitrons. De telles stratégies pourraient restaurer le splicing normal ou éliminer sélectivement des isoformes pathogènes, offrant une approche de médecine de précision pour le traitement du cancer. Le potentiel de cibler les mécanismes de splicing, y compris le splicing des exitrons, est activement exploré par des institutions de recherche et des entreprises pharmaceutiques du monde entier, avec plusieurs essais cliniques en cours pour des thérapies modulatrices du splicing (National Cancer Institute).
Au-delà de l’oncologie, le splicing des exitrons est impliqué dans une gamme de troubles génétiques et neurodégénératifs. La dérégulation du splicing des exitrons peut perturber la fonction normale des protéines, contribuant ainsi à des phénotypes de maladie. Les interventions thérapeutiques qui corrigent ou compensent ces défauts de splicing sont à l’étude, tirant parti des avancées dans les thérapies à base d’ARN et les technologies de modification génique. Par exemple, des approches basées sur CRISPR/Cas pourraient être utilisées pour modifier les éléments régulateurs du splicing, influençant ainsi l’inclusion ou l’exclusion des exitrons de manière contrôlée (National Institutes of Health).
En biotechnologie, la nature programmable du splicing des exitrons offre des outils pour la biologie synthétique et l’ingénierie des protéines. En concevant des gènes synthétiques avec des exitrons conçus, les chercheurs peuvent créer des protéines avec des domaines ou des caractéristiques régulatrices personnalisables, élargissant le répertoire fonctionnel des systèmes biologiques. Cela a des applications dans le développement de nouvelles enzymes, de biosenseurs et de protéines thérapeutiques. De plus, comprendre le splicing des exitrons améliore l’annotation des transcriptomes et des protéomes, améliorant ainsi la précision des modèles de gènes et des prédictions fonctionnelles tant dans la recherche fondamentale que dans la recherche appliquée (European Bioinformatics Institute).
Dans l’ensemble, l’élucidation des mécanismes du splicing des exitrons est prête à transformer les stratégies thérapeutiques et l’innovation biotechnologique, soulignant l’importance de la recherche continue et de la collaboration entre les acteurs académiques, cliniques et industriels.
Orientations Futures et Questions Ouvertes
Le splicing des exitrons, une forme récemment caractérisée de splicing alternatif où des régions exoniques internes (exitrons) sont excisées des ARN matures, a rapidement émergé comme un mécanisme significatif pour élargir la diversité transcriptomique et protéomique. Malgré les avancées dans son identification et son annotation fonctionnelle, plusieurs orientations futures et questions ouvertes demeurent critiques pour une compréhension complète de ses implications biologiques et cliniques.
Une grande area pour des recherches futures est l’élucidation des mécanismes régulateurs gouvernant le splicing des exitrons. Alors que le splicing canonique est orchestré par des composants et des facteurs régulateurs du spliceosome bien caractérisés, les éléments cis spécifiques et les protéines trans-agissantes qui déterminent la reconnaissance et l’excision des exitrons ne sont pas encore complètement définis. Des études de mutagenèse à haut débit et d’études de crosslinking, combinées avec des modélisations computationnelles avancées, sont nécessaires pour cartographier ces réseaux régulateurs. De plus, l’interaction entre le splicing des exitrons et d’autres événements de traitement de l’ARN, tels que l’édition de l’ARN et la polyadénylation alternative, reste largement inexplorée.
Une autre question ouverte concerne la conservation évolutive et la signification fonctionnelle du splicing des exitrons à travers les espèces. Des études initiales ont identifié le splicing des exitrons chez à la fois les plantes et les animaux, suggérant un mécanisme ancien et possiblement conservé. Cependant, l’ampleur à laquelle le splicing des exitrons contribue à la complexité des organismes, à l’adaptation, ou à la susceptibilité aux maladies n’est pas bien comprise. Des analyses de génomique comparative et des essais fonctionnels dans des organismes modèles divers seront essentiels pour aborder ces questions.
La pertinence clinique du splicing des exitrons est un domaine prometteur mais sous-développé. Des découvertes récentes indiquent que le splicing des exitrons peut générer des isoformes protéiques nouvelles avec des fonctions altérées, dont certaines pourraient agir en tant que néo-antigènes dans le cancer ou contribuer à la résistance aux médicaments. Cependant, la prévalence et l’impact des isoformes dérivées des exitrons dans les maladies humaines restent à être systématiquement caractérisés. Des analyses transcriptomiques à grande échelle d’échantillons de patients, couplées à une validation protéomique, sont nécessaires pour évaluer leur potentiel diagnostique et thérapeutique. De plus, le développement d’inhibiteurs ou de modulateurs spécifiques du splicing des exitrons pourrait ouvrir de nouvelles avenues pour des thérapies ciblées.
Enfin, les défis techniques associés à la détection et la quantification précises des événements de splicing des exitrons doivent être abordés. Les technologies de séquençage RNA actuelles et les pipelines bioinformatiques peuvent sous-estimer ou mal classer les événements des exitrons en raison de leurs sites de splice non canoniques et de leur longueur variable. Une innovation continue dans le séquençage à longues lectures et des outils d’annotation basés sur l’apprentissage machine sera cruciale pour faire progresser le domaine.
À mesure que la recherche progresse, les efforts collaboratifs entre les institutions académiques, les centres cliniques, et les consortiums internationaux tels que les National Institutes of Health et l’European Bioinformatics Institute seront essentiels pour standardiser les méthodologies et partager les données. Répondre à ces questions ouvertes ne pourra qu’approfondir notre compréhension de la biologie de l’ARN mais pourrait également révéler de nouvelles stratégies pour le diagnostic et le traitement des maladies.
Sources & Références
- National Institutes of Health
- European Bioinformatics Institute
- National Center for Biotechnology Information
- GENCODE
- Oxford Nanopore Technologies
- National Cancer Institute
- European Molecular Biology Laboratory